
 Hot off the press — the latest anole journal cover! In this issue of Nature Ecology & Evolution, Shane Campbell-Staton and I led a team of researchers to explore the effects of urban heat islands on anoles. We found that not only can urban Anolis cristatellus tolerate higher temperatures than their forest counterparts, but also identified genomic regions associated with divergent thermal tolerance. Check out a summary of this work at the urban evolution blog I co-edit, Life in the City: Anoles Adapt to Beat the Urban Heat.
Hot off the press — the latest anole journal cover! In this issue of Nature Ecology & Evolution, Shane Campbell-Staton and I led a team of researchers to explore the effects of urban heat islands on anoles. We found that not only can urban Anolis cristatellus tolerate higher temperatures than their forest counterparts, but also identified genomic regions associated with divergent thermal tolerance. Check out a summary of this work at the urban evolution blog I co-edit, Life in the City: Anoles Adapt to Beat the Urban Heat.
Tag: Anolis cristatellus Page 1 of 3

(Baeckens et al. 2019)
The skin surface structure of lizards varies greatly among species, likely because it plays a key role in a range of tasks, such as camouflage, locomotion, self-cleaning, mitigation of water loss and protection from physical damage. Yet, we still know remarkably little about how variation in skin surface structure translates to functional variation. Part of this gap in our understanding can be traced back to the lack of means to perform high-throughput and detailed analysis of the 3D anatomy of lizard skin in a non-destructive manner.
To tackle this hiatus, I was fortunate enough to be able to round up a great team of scientists and to start exploring the possibilities of a new imaging technique, termed gel-based stereo-profilometry. In this approach, a deformable transparent gel pad with one opaque surface is pressed onto the object of interest, creating a surface impression. While the gel pad is still in contact with the object, a series of photographs from six different illumination angles are acquired, and a topographical 3D map of the surface is created by merging the acquired images using
specialized surface analysis software.
Using this technique, we successfully imaged the 3D skin surface structure of Anolis cristatellus specimens in great detail (pixel resolution of 0.86 µm) and in a short-time frame (average acquisition time of imaging and digital reconstruction combined was 90 seconds). In our new paper, we demonstrate that this technique is exceptionally useful for the rapid 3D structural characterisation of lizard skin surfaces without any specimen preparation, permitting 3D visualization in situ and even in vivo. This technique opens exciting new avenues for investigating structure–function relationships in lizard skin.
In addition to the ability to quantify the micro- and macro-structural details of lizard skin, the 3D data sets acquired using gel-based stereo-profilometry can be directly converted into surface meshes, which can in turn be 3D printed. These tangible models can then be directly employed for studies to investigate the role of scale geometry on animal–substrate interactions, or enlarged for educational purposes to illustrate key differences between different squamate taxa.

Baeckens S, Wainwright DK, Weaver JC, Irschick DJ & Losos JB (2019) Ontogenetic scaling patterns of lizard skin surface structure as revealed by gel‐based stereo‐profilometry. Journal of Anatomy 235, 346–356.

Somebody needs to work on their anole species identification skills.
Breeding season is heating up for anoles in Miami, and at least one male crested anole (A. cristatellus) is a little…confused. While collecting some baseline data for my post-doc work looking at impacts of artificial light at night (ALAN) on brown and crested anoles, I noticed a commotion on a nearby cycad. Upon closer inspection, I realized that a male crested anole was pursuing and subsequently mating with a female brown anole (A. sagrei) who was decidedly unhappy about the situation.
In case you’re wondering about the colorful jewelry at the base of their tails, both of the anoles in the photo/video are bead-tagged to allow me to reidentify them from a distance. The copulation here lasted 3-4 minutes a portion of which I managed to capture on video.
While previous reports on AA have documented coupling between A. carolinensis and A. sagrei, I haven’t seen any reports of interspecific mating between A. cristatellus and A. sagrei. Has anyone else observed this phenomenon? The two species do encounter each other quite frequently in the Miami area, so this might not be a rare occurrence. Hybridization seems unlikely given the divergence between these two species, but you never know!
Lead author, Kirsten Tyler, reports on her recent Journal of Herpetology paper with K. Winchell and L. Revell:
Urbanization creates drastic changes to habitats leading to differences in microclimate, perch characteristics and distribution, and ecological communities (competitors, prey, and predators) when compared to natural (forest) habitats. Studies have found increased rates of mortality of many urban species due to generalist urban-tolerant predators such as raccoons, feral cats, and domestic animals (Ditchkoff 2006). Anolis lizards are able to voluntarily drop their tails (“autotomize”) when challenged by a predator, enabling their escape in many instances. The maimed lizards are able to regenerate their lost tails, though the replacement tail is a rod of cartilage and not the original bony vertebrae. The regenerated tail portions are often a different color and texture, and the lack of vertebrae / cartilage rod are clearly visible in X-rays.

A. cristatellus with an autotomized tail in a forest habitat

A. cristatellus with an autotomized tail in an urban habitat
We hypothesized that autotomy rates would be more similar between urban areas in different municipalities than to natural areas in the same municipality due to similar predator regimes in urban sites across the island. We compared the frequency and pattern (number of caudal vertebrae remaining) of caudal autotomy of A. cristatellus between urban and natural areas in Puerto Rico.

X-rays of our samples with an intact tail (A) and an autotomized tail (B).
We sampled A. cristatellus from paired natural and urban sites in four Puerto Rican municipalities: San Juan, Mayagüez, Ponce, and Arecibo. The natural sites were high quality natural forests and the urban sites were high-density residential areas. Urban sites were dominated by asphalt and other impervious surfaces, had sparse tree cover, and a large fraction of potential perches were manmade surfaces such as walls and fences. We scored 967 X-rays from these eight sites for caudal autotomy and counted the number of remaining tail vertebrae. We tested for an effect of urbanization on caudal autotomy by fitting a logistic regression model with municipality (San Juan, Mayagüez, Ponce, Arecibo) and site type (urban, natural), and their interactions, as model factors, and body size as a covariate.

Our data shows that lizards found in urban sites have a larger probability of having autotomized tails.
Interestingly, we found higher rates of autotomy in all urban populations compared to nearby natural areas. Differences in autotomy might be explained by differences in predator density and efficiency (Bateman 2011). For example, inefficient predators (those that more often than not fail to capture their prey) tend to leave behind more lizards with broken and regenerated tails (Schoener 1979). In addition, a greater abundance of predators could result in more predation attempts. Unfortunately, we did not collect data on predator abundances or community composition, so we cannot distinguish between these (non-mutually exclusive) explanations. Higher rates of autotomy in urban areas could thus reflect any of a variety of factors, including (but not restricted to) inefficient predators in urban areas, a shortage of refuges offering protection from predators, or an increase in predator density.

For lizards with autotomized tails, we found no significant difference in caudal vertebrae number between urban and natural sites.
Lastly, we did not find that lizards with autotomized tails in urban areas had lost more (or less) of their original tail to caudal autotomy. Since regenerated tails cannot be autotomized past the original break point (i.e. cartilage cannot autotomize), this suggests that lizards in urban areas are no more likely to be subject to multiple unsuccessful predation attempts (resulting in caudal autotomy) than lizards in natural forest. Future investigation quantifying predation attempts or predator community composition in urban and forest habitats could help us better understand the source of this intriguing pattern.
References:
BATEMAN, P. W., AND P. A. FLEMING. 2011. Frequency of tail loss reflects variation in predation levels, predator efficiency, and the behaviour of three populations of brown anoles. Biological Journal of the Linnean Society 103:648–656.
DITCHKOFF, S. T. 2006. Animal behavior in urban ecosystems: modifica- tions due to human-induced stress. Urban Ecosystems 9:5–12.
SCHOENER, T. W. 1979. Inferring the properties of predation and other injury-producing agents from injury frequencies. Ecology 60:1110–1115.

Kevin Aviles-Rodriguez, from the Revell lab at U. Mass. Boston, gave the second urban anole-themed talk of the meeting. Kevin presented his Master’s thesis work that he conducted with the Kolbe lab at U. Rhode Island in a talk titled, “Structural habitat alterations caused by urbanization influence escape behavior of a common lizard.”
Urban habitats are drastically modified and present novel resources and threats for animals that persist and utilize these spaces. Structurally, urban habitats have different types of surfaces that are smoother, broader in diameter, and often more vertically oriented (90° angle). Urban habitats also present abundant and novel food resources in terms of human food and insects attracted to lights and garbage. But with the abundance of food and novel niche space also comes an abundance of novel predators such as cats and dogs kept as pets.
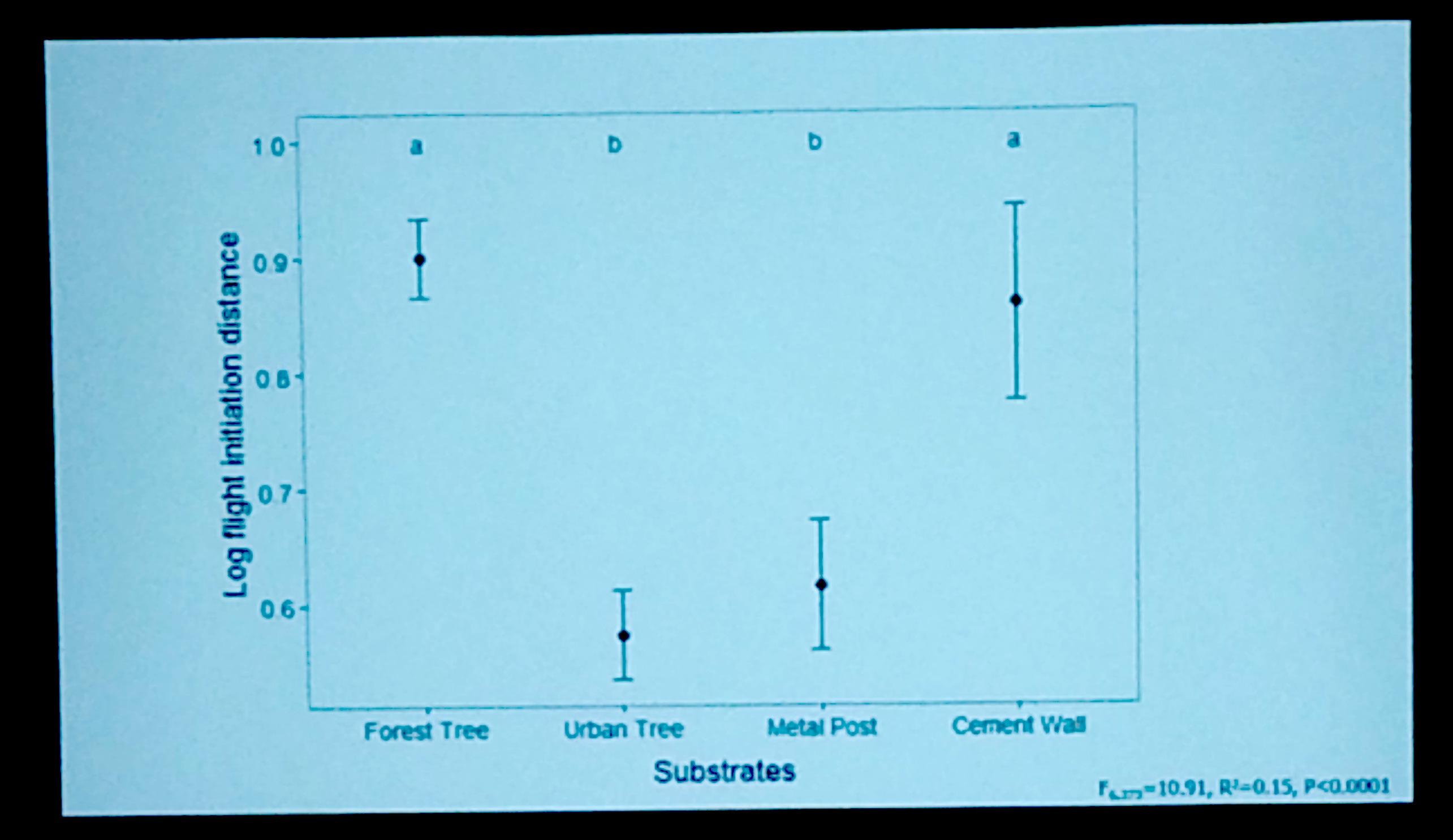
Kevin wanted to know how Anolis cristatellus from San Juan, Puerto Rico and South Miami behaved in urban habitats compared to forest habitats when perceiving a predation threat. Although there are obvious costs of not escaping a predator successfully, there are also costs of fleeing when not necessary in terms of lost feeding opportunities and disrupted social interactions (mating, territory defense). Kevin wanted to know if the urban environment influenced escape behavior decisions. Specifically, he had two objectives: (1) To quantify escape behavior (squirreling, jumping, or sprinting) and how this relates to different types of perches found in urban areas. (2) To measure flight-initiation distance (FID), or how close one can approach an animal before it flees, to see if there are differences between forest lizards and urban lizards.
 Kevin found that as perch diameter increases, the probability that a lizard will squirrel around a perch or sprint up the perch increased and the probability of jumping decreased. Interestingly, when he also looked at perch use, he found that the majority of lizards were using perches of thinner diameter where the probability of jumping was highest. Urban lizards also tended to use more isolated perches, which he defined as the number of nearby potential perches within 1 meter. When nearby perch density was lower, lizards tended to jump less – perhaps not all that surprising since they have fewer places to jump to. Kevin also found that escape strategy differed based on the type of perch used. In urban habitats, on trees and on metal posts lizards squirreled more frequently than they did in forest habitats. Interestingly, on cement walls (e.g. buildings) lizards did not jump at all and mainly sprinted to escape.
Kevin found that as perch diameter increases, the probability that a lizard will squirrel around a perch or sprint up the perch increased and the probability of jumping decreased. Interestingly, when he also looked at perch use, he found that the majority of lizards were using perches of thinner diameter where the probability of jumping was highest. Urban lizards also tended to use more isolated perches, which he defined as the number of nearby potential perches within 1 meter. When nearby perch density was lower, lizards tended to jump less – perhaps not all that surprising since they have fewer places to jump to. Kevin also found that escape strategy differed based on the type of perch used. In urban habitats, on trees and on metal posts lizards squirreled more frequently than they did in forest habitats. Interestingly, on cement walls (e.g. buildings) lizards did not jump at all and mainly sprinted to escape.  Kevin offered a few possible explanations for this trend. For one, building perches tend to be more isolated than trees and so it may simply be that lizards on these substrates have nowhere to jump to. A second possibility is that the lizards have trouble jumping from these perches since they are more vertical than the optimal angle for jumping (39-42°, Toro et al. 2003).
Kevin offered a few possible explanations for this trend. For one, building perches tend to be more isolated than trees and so it may simply be that lizards on these substrates have nowhere to jump to. A second possibility is that the lizards have trouble jumping from these perches since they are more vertical than the optimal angle for jumping (39-42°, Toro et al. 2003).
In his final analysis, Kevin found that flight initiation distance (how close you can get to the animal before it flees) was very short for animals perched on urban trees and metal posts. In fact, he commented that on some occasions he was able to get close enough to touch the lizard before it fled! This difference was significantly shorter than for animals perched on trees in the forest and for animals perched on painted concrete walls in the city.
{kind=link}

Quynh Quach presenting her Master’s thesis work at JMIH.
Quynh Quach, a master’s student from the Revell Lab at U. Mass. Boston, presented her thesis research on “Phylogeography and Population Structure of Anolis cristatellus on the island of Vieques.” Before Quynh joined the Revell lab, former post-doc Graham Reynolds and former Losos lab undergraduate Tanner Strickland looked at the phylogeography of Anolis cristatellus across Puerto Rico and the Virgin Islands using mitochondrial DNA (in review). Tanner’s work revealed that there was a mitochondrial break on the island of Vieques, just off the coast of Puerto Rico. The mitochondrial data suggested that there were two genetically different groups of A. cristatellus, one on the East and one on the West of Vieques. The only problem was, as we know, mtDNA patterns are not always supported by nuclear whole-genome DNA patterns. In addition, Tanner’s dataset only consisted of 9 samples from Vieques.
When Quynh joined the lab, she wanted to know more about this pattern. Would this division be supported by nuclear genome analyses? Were these lineages anthropogenically introduced? If not, what was the origin of these groups – historical allopatry followed by secondary contact or isolation by distance? So she set out to answer these questions by collecting 300 tail tips from across the island of Vieques, extracting and sequencing both mtDNA and nuclear DNA.

The mtDNA variation shows a strong geographic pattern.
Quynh first constructed a mitochondrial phylogeny to verify the pattern observed by Tanner and Graham. The mtDNA analysis confirmed that there are 2 mtDNA clades on Vieques with strong geographic patterns. The island-wide pattern of mtDNA variation was not what we would expect if anthropogenic introduction were the cause since this would be unlikely to show such a clear East-West pattern with the small contact zone in the middle. So then how did this pattern arise?

Analysis with K=2 shows two clear groups associated with the East and West.
To answer that question, Quynh next looked at nuclear DNA using RADseq. She sequenced 48 individuals: 5 from Virgin Islands, 6 from Puerto Rico, and 37 from Vieques, then de novo assembled the genome and called 16,808 SNP’s. She ran STRUCTURE and DAPC analyses on this data and found that the Virgin Island samples form 1 cluster and Puerto Rico and Vieques form a second cluster with 4.1% divergence between the groups. But she wondered, what if we look at just Vieques and specify K=2? When she did this with DAPC and saw a clear geographic pattern similar to what she found with the mtDNA. Finally, she tested whether this represented isolation by distance. She found that there was significantly reduced gene flow between geographically distant individuals, supporting this hypothesis as the most likely cause of the variation.
Lastly, Quynh emphasized that it is important to consider multiple genetic markers and not just rely on mtDNA results. Had the group stopped at their original mitochondrial analysis, they would have reached a very different conclusion.

Anolis cristatellus on a smooth, vertical substrate in Puerto Rico (photo by K. Winchell)
In urban areas, the number of natural substrates (e.g. trees) is reduced. In their place are novel manmade substrates (e.g. walls, metal gates). These surfaces undoubtedly have different properties relevant to anole locomotion: they are smoother, harder, and (in the case of walls) much broader and flatter compared to natural surfaces in a forest. In urban areas lizards still use these substrates at high frequency, but do they do so effectively? Kolbe and co-authors began to dive into this complex topic in their recent publication, “City slickers: poor performance does not deter Anolis lizards from using artificial substrates in human-modified habitats” (Kolbe et al. 2015).
The relationship between habitat use, morphology, and performance for anoles has been extensively studied in natural environments (reviewed in Losos 2009). Urban environments add new dimensions to this area of research. Resource distribution and abundances differ drastically compared to natural areas. For example, the distribution of available perches and what they are made of in urban habitats is very different from a forest. Moreover, the properties of these resources differ drastically as well: urban substrates are smoother, broader, and have different thermal properties, to start. Understanding these differences in habitat use and how they influence performance and, ultimately, adaptive responses in anoles is the topic of ongoing collaborative research that I (K. Winchell) and the Kolbe lab have been conducting.
With a flurry of recent attention investigating how background light may influence the signalling efficiency of Anolis dewlaps (1,2,3,4), particularly those inhabiting low-light environments where patches of sunlight appear at a premium, it occurred to me that extended dorsal and tail crests may fall under similar selection. Below are some photos of Puerto Rican crested anoles (Anolis cristatellus) – a species in which males exhibit an enlarged tail crest and the ability to voluntarily erect impressive nuchal and dorsal crests during aggressive interactions (the mechanisms of which are detailed in this previous AA post) – that show how crests may contribute to signalling.

I have no doubt this thought has crossed the minds of many anole scientists before, particularly those current graduate students so successfully studying A. cristatellus and familiar with their ecology and behaviour (namely Alex Gunderson, Kristin Winchell, Matt McElroy, and Luisa Otero). Dewlaps are undoubtedly of primary importance to anole signalling and communication, but what are people’s general thoughts on the relative importance of other morphological features?


An actual perch used by Anolis cristatellus – neither smooth nor flat! (photo by K. Winchell)
In the real world, lizards cling to everything from smooth, flat concrete walls to rough, rounded tree trunks. So why is it that most studies on cling force in anoles focus on clinging to smooth flat substrates? Does cling force differ if the substrate is rounded or rough? Jason Kolbe sought to answer this question in his recent publication, “Effects of Hind-Limb Length and Perch Diameter on Clinging Performance in Anolis Lizards from the British Virgin Islands” (Kolbe 2015).
We know that morphology impacts performance in anoles and that performance varies with environment. For example, sprint speed is correlated with limb length, but this relationship depends on the diameter of the substrate (e.g. Losos and Sinervo, 1989). We also know a little about clinging performance in anoles. Greater cling force is correlated with larger toepads and more lamellae on smooth flat surfaces (Irschick et al., 1996; Zani 2000; Elstrott and Irschick, 2004), but adhesion on rougher surfaces may be influenced by claw and toe morphology (Zani 2000).
There appears to be an unexplored interaction between substrate properties and clinging ability that involves more than just toepad characteristics. Specifically, Kolbe points out that claws can increase clinging ability by digging into the perch or simply by increasing friction on the surface. Limbs can also increase friction via the application of compression forces to the substrate. In other words, cling force can be increased, particularly on rough surfaces, by using muscular force to grasp rather than relying on van der Waals forces from the toepads, which are more effective on smooth flat surfaces.

Anolis cristatellus (photo by KMW)

Anolis stratus (photo by KMW)
Anole species used in this study: Anolis cristatellus (left) and Anolis stratulus (right). Photos by K. Winchell.
Kolbe investigated this further by looking at the interaction between limb length and clinging ability on perches of different diameters with Anolis cristatellus and Anolis stratulus from the British Virgin Islands. Specifically, he hypothesized that cling force should increase as the ability of a lizard to obtain a firm grasp on a substrate improves (i.e. when it can wrap its limbs around the substrate). This ability to form a secure grasp is dependent on both the diameter of the perch and on lizard limb length.

Anolis cristatellus in survey posture (photo by K. Winchell)
Foraging decisions are the result of a complex decision-making process involving intrinsic factors (physiology, body condition, cognitive ability, sex, ontogeny, etc.) and environmental factors (food availability, structural habitat, presence of predators and competitors). In short, it comes down to the tradeoff between the benefits of energetic gain and the potential costs of predation risk, missed opportunities for reproduction, and expended energy. However, little is known about the specifics of this process – what information are lizards considering when making this decision? By conducting manipulative field experiments on Anolis cristatellus in Puerto Rico, Drakeley et al. (2015) attempt to elucidate what environmental factors influence the decision to forage.
The authors conducted field experiments involving feeding trays in the wild. The Puerto Rican crested anole is a trunk-ground anole and a sit-and-wait forager. When receptive to feeding, it perches head down in “survey posture,” a behavior it reduces when satiated. Aside from movement associated with foraging and social interactions, this species typically remains stationary on a perch. Because of this, the authors were able to easily locate a focal individual and count the number of conspecifics present, using natural variation instead of manipulating the number of animals present.
In the first experiment, they manipulated the food quantity to determine how foraging decisions differ when food is plentiful versus scarce and how this is influenced by the presence of competitors. They found that lizards foraged faster when there were more conspecifics present and food was scarce. When no lizards were near the feeding tray and the feeding tray was full, the focal animal took longer to approach the tray to take the mealworms compared to when there were many conspecifics nearby. Interestingly, this was not related to overall local density, but rather to the number of conspecifics in the immediate vicinity. Therefore the decision to forage likely involves an instantaneous assessment of the local conditions rather than knowledge of the long-term population trends. The authors also considered several other factors and found that although body size was related to foraging latency (larger lizards were quicker to the feeding tray), no other environmental factors were relevant (temperature, humidity, perch height, perch diameter, local density of conspecifics).

Figure 1 from Drakeley et al. (2015). Latency to feed was correlated with the number of conspecifics present and abundance of food.
In the second experiment, the authors chose focal animals farther from the feeding trays and considered distance as a proxy for predation risk. The farther the lizard was from the tray, presumably the greater exposure it had to predators as it moved towards the tray. They found that under this scenario, when risk was elevated, there was more latency in the approach of the food tray. This effect was driven mainly by the increased use of intermediate perches rather than a direct approach across open ground. Increased latency to feed was observed regardless of how abundant the food was or how many conspecifics approached the tray, supporting the conclusion that this effect was because of the perception of greater predation risk (i.e. movement over a longer distance). They also found that larger lizards had a lower latency to feed (approached the feeding tray more rapidly) and lizards not in the foraging position had a longer latency to feed.
In summary, it seems that anole foraging decisions are quite complex. Lizards appear to weigh the risk of predation taking cues from conspecific behavior and abundance versus the abundance of food to make instantaneous decisions to approach a novel feeding source.
Drakeley M, Lapiedra O, Kolbe JJ (2015) Predation Risk Perception, Food Density and Conspecific Cues Shape Foraging Decisions in a Tropical Lizard. PLoS ONE 10(9): e0138016. doi:10.1371/journal.pone.0138016