
Kevin Aviles-Rodriguez, from the Revell lab at U. Mass. Boston, gave the second urban anole-themed talk of the meeting. Kevin presented his Master’s thesis work that he conducted with the Kolbe lab at U. Rhode Island in a talk titled, “Structural habitat alterations caused by urbanization influence escape behavior of a common lizard.”
Urban habitats are drastically modified and present novel resources and threats for animals that persist and utilize these spaces. Structurally, urban habitats have different types of surfaces that are smoother, broader in diameter, and often more vertically oriented (90° angle). Urban habitats also present abundant and novel food resources in terms of human food and insects attracted to lights and garbage. But with the abundance of food and novel niche space also comes an abundance of novel predators such as cats and dogs kept as pets.
Kevin wanted to know how Anolis cristatellus from San Juan, Puerto Rico and South Miami behaved in urban habitats compared to forest habitats when perceiving a predation threat. Although there are obvious costs of not escaping a predator successfully, there are also costs of fleeing when not necessary in terms of lost feeding opportunities and disrupted social interactions (mating, territory defense). Kevin wanted to know if the urban environment influenced escape behavior decisions. Specifically, he had two objectives: (1) To quantify escape behavior (squirreling, jumping, or sprinting) and how this relates to different types of perches found in urban areas. (2) To measure flight-initiation distance (FID), or how close one can approach an animal before it flees, to see if there are differences between forest lizards and urban lizards.
 Kevin found that as perch diameter increases, the probability that a lizard will squirrel around a perch or sprint up the perch increased and the probability of jumping decreased. Interestingly, when he also looked at perch use, he found that the majority of lizards were using perches of thinner diameter where the probability of jumping was highest. Urban lizards also tended to use more isolated perches, which he defined as the number of nearby potential perches within 1 meter. When nearby perch density was lower, lizards tended to jump less – perhaps not all that surprising since they have fewer places to jump to. Kevin also found that escape strategy differed based on the type of perch used. In urban habitats, on trees and on metal posts lizards squirreled more frequently than they did in forest habitats. Interestingly, on cement walls (e.g. buildings) lizards did not jump at all and mainly sprinted to escape.
Kevin found that as perch diameter increases, the probability that a lizard will squirrel around a perch or sprint up the perch increased and the probability of jumping decreased. Interestingly, when he also looked at perch use, he found that the majority of lizards were using perches of thinner diameter where the probability of jumping was highest. Urban lizards also tended to use more isolated perches, which he defined as the number of nearby potential perches within 1 meter. When nearby perch density was lower, lizards tended to jump less – perhaps not all that surprising since they have fewer places to jump to. Kevin also found that escape strategy differed based on the type of perch used. In urban habitats, on trees and on metal posts lizards squirreled more frequently than they did in forest habitats. Interestingly, on cement walls (e.g. buildings) lizards did not jump at all and mainly sprinted to escape.  Kevin offered a few possible explanations for this trend. For one, building perches tend to be more isolated than trees and so it may simply be that lizards on these substrates have nowhere to jump to. A second possibility is that the lizards have trouble jumping from these perches since they are more vertical than the optimal angle for jumping (39-42°, Toro et al. 2003).
Kevin offered a few possible explanations for this trend. For one, building perches tend to be more isolated than trees and so it may simply be that lizards on these substrates have nowhere to jump to. A second possibility is that the lizards have trouble jumping from these perches since they are more vertical than the optimal angle for jumping (39-42°, Toro et al. 2003).
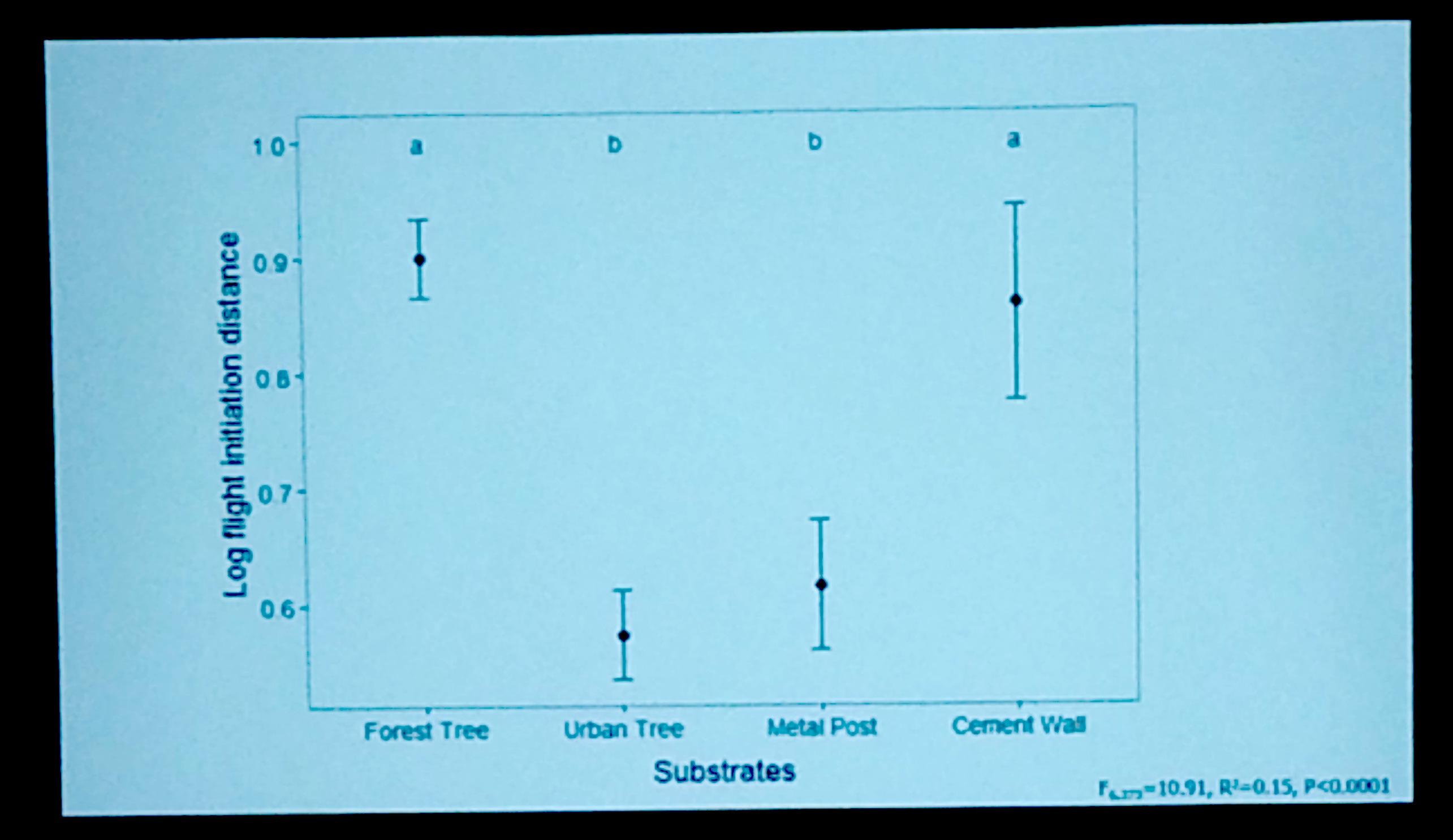
In his final analysis, Kevin found that flight initiation distance (how close you can get to the animal before it flees) was very short for animals perched on urban trees and metal posts. In fact, he commented that on some occasions he was able to get close enough to touch the lizard before it fled! This difference was significantly shorter than for animals perched on trees in the forest and for animals perched on painted concrete walls in the city.


































{kind=link}